MODULE 1 - VIEWS-ALL 1
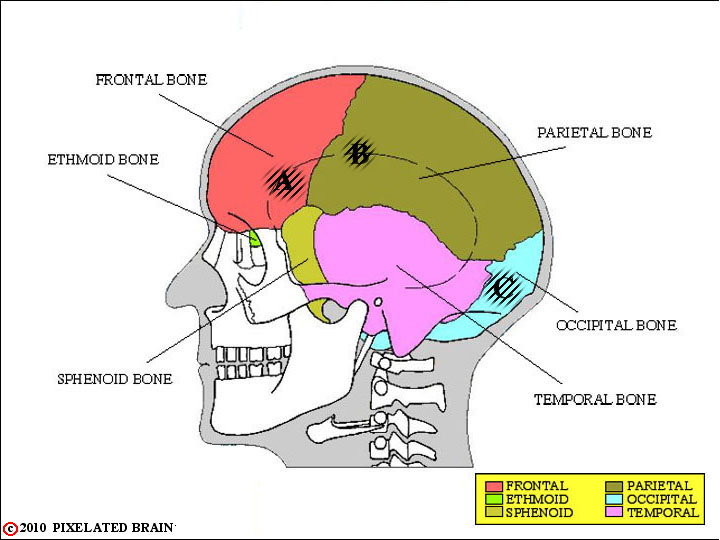
FIGURE 1-1
This lateral view simply shows the 6 bones that contribute to the surface of the cranial cavity. Of course, what we really care about is the relationship of these bones to the brain. If a penetrating wound passed through a given part of the temporal bone, for example, what region of the brain would it damage? To get some idea of the answer, go on to Fig. 1-2.
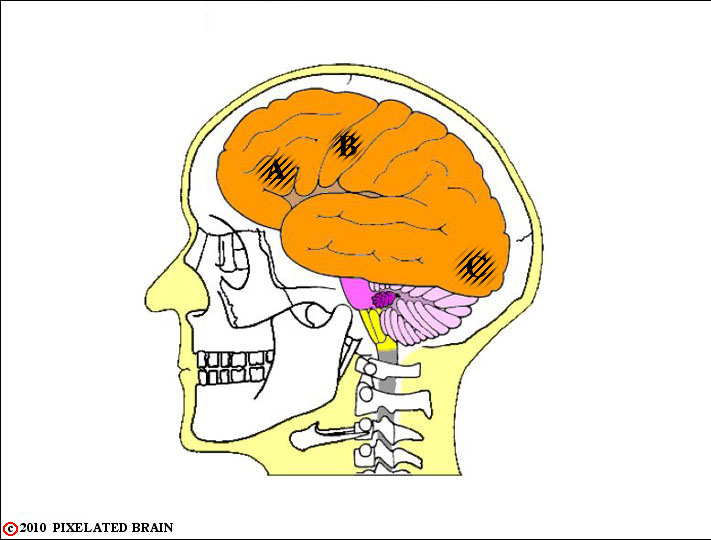
FIGURE 1-2
Now, we look right through the skull to get a lateral view of the brain, within the cranial cavity.

FIGURE 1-3
Now, we look right through the skull to get a lateral view of the brain, within the cranial cavity. .

FIGURE 1-4
In this view we look down at the floor of the cranial cavity. On the right, some of the obvious landmarks are labeled. On the left, we have tried to introduce you to the concept of the three cranial fossae. These terms are frequently used, clinically, to describe different regions within the cranial cavity.

FIGURE 1-5
This view simply shows the major openings in the skull. When we take up the cranial nerves, later on, you will want to be able to say which nerves or structures pass through each of these openings.

FIGURE 1-6
Certainly, the most important function of the skull is to protect the fragile brain, lying within the cranial cavity. As this view indicates, the skull is well-designed for the job. If we cut through the calvarium in the plane shown in A and look down from above we get view B. The contour of the brain's surface matches that of the cranial cavity rather well, but there is a small space between the two. This narrow gap, shown by green stippling in the figure, is called the subarachnoid space. It is filled with cerebrospinal fluid (CSF) and also contains three membranes known collectively as the meninges. The brain is, in a sense, floating in CSF which protects it from "bumping into" the skull during sudden movements of the head. From a clinical point of view, the anatomy of this region is extremely important and we want to look at it in detail. The best way to start is to consider CSF - where does this fluid come from, how does it get into the subarachnoid space, where is it going, and how does all this relate to the meninges? To come up with the answers, we will jump around a bit. First, let's look at a few views taken from an old lecture on brain development.

DEV-1
In the first few days following fertilization of the ovum, the single celled zygote divides repeatedly to form a solid cell mass of 12 - 16 cells called a morula. During the second week of development this structure implants in the wall of the uterus. Two cavities now develop within the mass - the amnionic cavity above and the yolk sac below. Between these two lies the embryonic bilaminar disc, consisting of a layer of ectoderm lying above a layer of entoderm. During the third week of development a region termed the primitive streak forms on the dorsal surface of the disc, giving a longitudinal axis to the developing embryo. Cells from the surface of the disc migrate through this region and then spread out between the ectoderm and the entoderm to create a new cell layer. The cells that migrate laterally form mesoderm, thus transforming the bilaminar disc into a trilaminar one. Other cells migrate anteriorly in the midline to form the notocord. By a process of neural transduction, the notocord induces the overlying ectoderm to become neural tissue - the neural plate. In the midline, just above the notocord, a neural groove forms. Lateral to this on either there is a proliferation of neural ectoderm to form neural folds.

FIGURE DEV-2
On about day 18 following fertilization, the process of neurulation begins. Starting in the cervical region, the neural folds migrate dorsally and fold toward the midline to meet and fuse, forming the first part of the neural tube. Fusion of the folds then extends both rostrally and caudally. For several days the ends of the neural tube remain open, so that the central canal is in communication with the amnionic cavity. The rostral opening, or cranial neuropore, is closed at about day 25 by a membrane called the lamina terminalis. The caudal neuropore closes a few days later.
Failure of the cranial neuropore to close leads to anencephaly, a developmental defect which grossly distorts the development of the brain. Failure of the caudal neuropore to close is associated with some form of spina bifida.
Some tissue of the neural plate fails to become encorporated in the neural tube and remains just dorsolateral to it. This neural crest tissue ultimately gives rise to many structures, including: 1) the dorsal root (sensory) ganglia of the peripheral nervous system; 2) the ganglia of the autonomic nervous system; 3) the cells of the adrenal medulla and 4) the Schwann Cells that form the myelin that surrounds peripheral nerves.
Lateral to the neural tube the mesoderm becomes organized in a segmental fashion into somites which will, ultimately, impose a segmental pattern upon the peripheral nervous system.

DEV-3
The rostral end of the neural tube undergoes a series of regional expansions to generate the final form of the brain. In early development (3-4 weeks) there are three primary vesicles:
1. Forebrain = Prosencephalon
2. Midbrain = Mesencephalon
3. Hindbrain = Rhombencephalon

DEV-3
The rostral end of the neural tube undergoes a series of regional expansions to generate the final form of the brain. In early development (3-4 weeks) there are three primary vesicles:
1. Forebrain = Prosencephalon
2. Midbrain = Mesencephalon
3. Hindbrain = Rhombencephalon
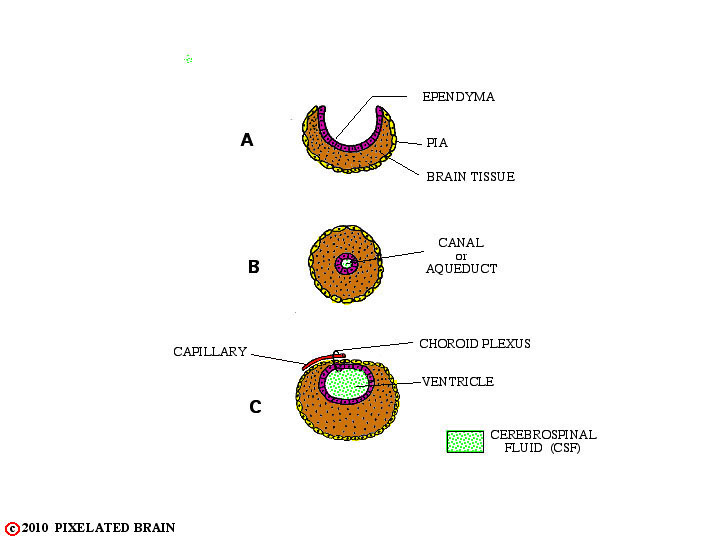
FIGURE 1-7
In two regions, however, the edges of the neural plate fail to meet, as shown A. In these instances , the ependymal cell layer, lining the cavity, and the layer of pial cells, covering the surface of the tube, meet to form a membrane which completes closure of the cavity. Blood vessels extend into this membrane creating highly vascular tissue called choroid plexus (see B and C, above). The two regions where this occurs are shown in the highly schematic Figure 8.
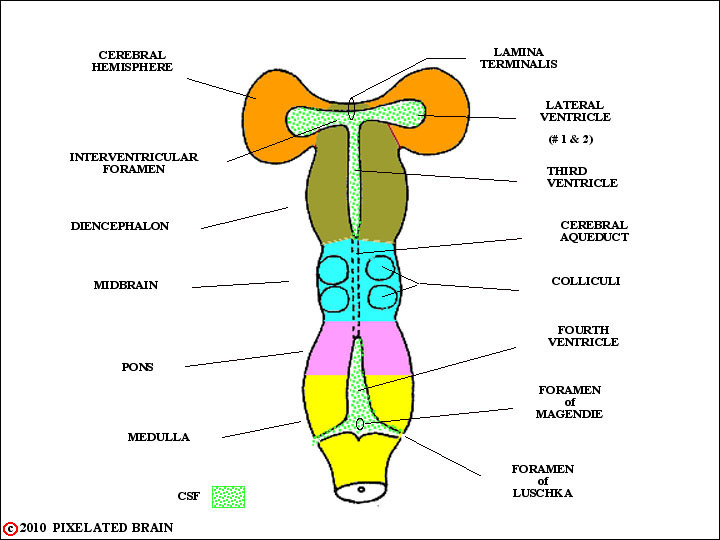
FIGURE 1-8
As development of the brain continues, regional variations in the wall of the tube become apparent. These regions, labeled on the left in this figure, will become the major subdivisions of the adult brain. Rostrally (i.e., at the very "top" of the neural tube) the lamina terminalis seals the ventricular system closed
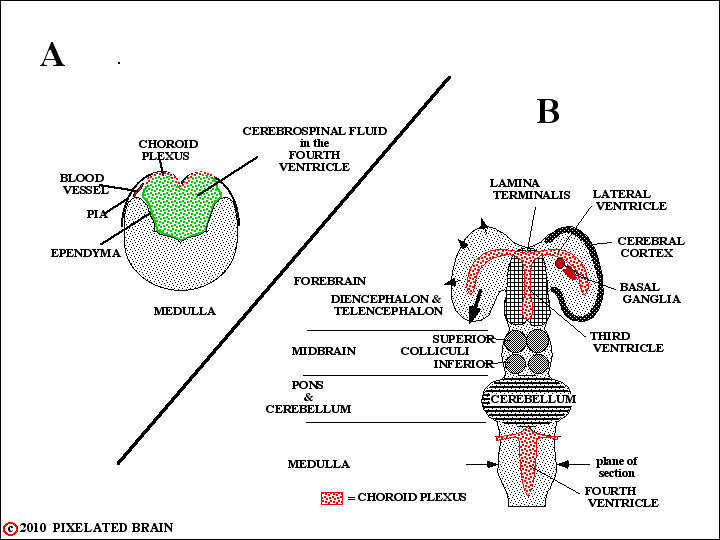
DEV-3
In some regions the edges of the neural plate fail to meet. In these instances , the ependymal cell layer, lining the cavity, and the layer of pial cells, covering the surface of the tube, meet to form a membrane which completes closure of the cavity. Blood vessels extend into this membrane creating highly vascular tissue called choroid plexus. The two regions where this normally occurs are shown in this highly schematic figure. In A, which is simply a restatement of Figure 7, we see the situation in the medulla. In B, which is a restatement of Figure 8, we see that the edges of the neural tube also fail to meet at the level of the diencephalon. Thus both the third and fourth ventricles are roofed by choroid plexus. Furthermore the choroid plexus of the third ventricle is carried out into the lateral ventricles within the hemisphere. Choroid plexus tissue is the main source of CSF, and since the greatest mass of this tissue is found lining the two lateral ventricles this is where most CSF is produced.

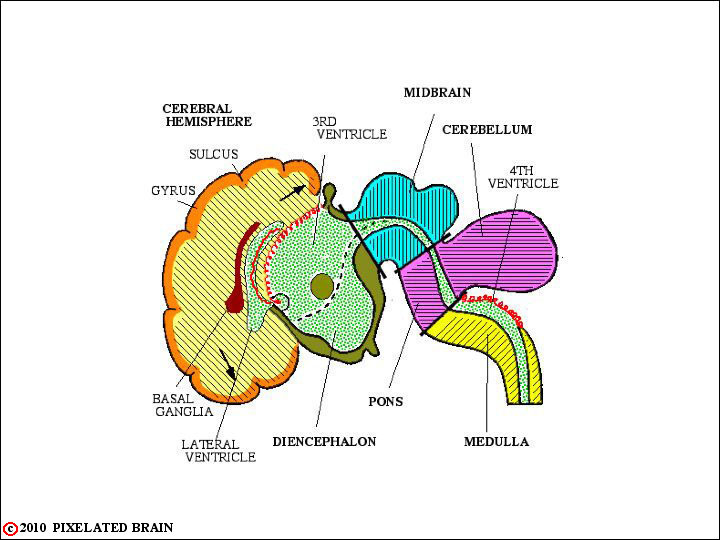
FIGURE 1-9
This viewtraces the flow of CSF. From the major site of formation, in the two lateral ventricle, fluid passes through the interventricular foramen to enter the third ventricle. A bit more fluid comes from the choroid plexus roofing the third ventricle; CSF exits into the cerebral aqueduct and descends into the fourth ventricle. Again, choroid plexus in the roof of this ventricle contributes a small amount of CSF to the total. CSF exits from the ventricular system through paired lateral openings, the Foramina of Luschka, and a single midline opening in the roof of the ventricle, the Foramen of Magendie. Once outside, the fluid lies within the subarachnoid space - a region defined more precisely in a coming view. For now, just trace the arrows up over the surface of the brain to the arachnoid granulation. CSF enters these specialized structures and from here returns to the venous blood of the superior sagittal sinus.
Fenestrated capillaries within choroid tissue allow for free passage of water and solutes but that tight junctions between choroid epithelial cells form a blood-CSF barrier. While lipid soluble substance may easily pass through this barrier, most other materials must be conveyed from blood to CSF by active transport through cells of the choroid epithelium. The result is that the chemical composition of the two fluids differ; for example, CSF has higher levels of chloride, magnesium and sodium, compared with plasma. CSF is colorless, low in protein and contains very few, if any, white blood cells.
The volume of CSF in the adult is about 120mL and roughly 500mL of the fluid is produced every day, so that about 4 turnovers occur in 24 hour. Production continues even if there is a blockage of flow, somewhere along the way to the arachnoid granulations, but the result depends on the age of the subject and the site of the obstruction. Most CSF returns to the vascular system by entering arachnoid granulations. Some fluid passes between the cells lining a granulation to mix with the venous blood of the superior sagittal sinus. Most of the CSF, however, is transported through the cells in membrane bound vesicles. Granulations are easily seen in gross specimens. If the dura is carefully removed from the surface of the brain, they tend to pop out of the superior sagittal sinus and be exposed to view, as in Figure 1-13. Another approach is to open the superior sagittal sinus in the midline and look at the granulations as they protrude into the sinus (Figure 1-14).

FIGURE 1-9a
This is a schematic section through the brain that will be useful in describing the flow of CSF and the meninges. Use it (as well as the other figures of this group), to confirm the following:
1) The dura may be thought of as a two layered structure, with an outer periostial layer (which serves as the periosteum of the skull) and an inner meningeal layer. The periostial layer is tightly adherent to the overlying bone, and in most regions the two layers of the dura are fused.
2) The two layers split to enclose the middle meningeal artery. The artery is positioned between the two layers in such a way that it creates a ridge on the outer (periostial) surface of the dura, but leaves the inner (meningeal) surface quite smooth (Figures. 1-7 & 1-8). One of the results of this arrangement is that the artery "erodes" a valley on the inner surface of the skull. Because skull fractures which cross this valley may well tear the artery and lead to intracranial bleeding, the precise course of the artery is of clinical significance.3) The two layers also split to enclose the venous sinuses of the cranial cavity. Three of these - the superior sagittal sinus, the inferior sagittal sinus and the straight sinus - are shown in Figure 1-9. Others you should be aware of include the transverse sinus, the sigmoid sinus and the superior and inferior petrosal sinuses.
4) The meningeal layer of the dura evaginates, or folds in upon itself, to create several partial partitions within the cranial cavity. These include the falx cerebri (Fig, 1-9), the falx cerebelli (Figure 1-10) and the tentorium cerebelli.
5) At the foramen magnum the two layers of dura separate (Figure 1-10); the endosteal layer passes around to the external surface of the skull while the meningeal layer descends within the spinal column to form a covering of the spinal cord.
6) The dura is sensitive to pain. Trigeminal fibers supply the dura of the anterior and middle cranial fossae, as well as the tentorium. The vagus nerve and cranial nerves 2 and 3 supply the dura of the posterior cranial fossa.
When the dura is lifted off the brain, the next meningeal layer, the arachnoid, is exposed. The two layers separate easily except in the region of the superior sagittal sinus. Here, two structures impede the separation: 1) "bridging veins", draining the surface of the hemisphere, pass through the arachnoid to empty into the sinus and 2) small evaginations of arachnoid, termed arachnoid granulations, push their way into the interior of the sinus. Arachnoid granulations represent the sites where cerebrospinal fluid, secreted into the ventricles of the brain, is returned to the venous system. We will return to the topic of cerebrospinal fluid later.

FIGURE 1-10
Clearly, there is an intimate relationship between CSF and the meninges so let's consider these tissues next. The outermost of the three layers is the dura - short for dura mater. It is a tough, fibrous layer, having the feeling and strength of canvas.To prepare this specimen, a large piece of dura was carefully pried away from the skull and left in place, covering the dorsal surface of the brain.

FIGURE 1-11
The dura consists of an outer periosteal layer, which is tightly adherent to the skull, and an inner meningeal layer. In most regions the two layers appear fused into a single one, and some textbooks describe it that way.
Deep to the dura lies the arachnoid, a delicate membrane that is easily separated from the dura in most places, but pressed tightly against it by the pressure of the CSF lying in contact with it's inner surface. The third meningeal layer is the pia, a fine cellular layer which is adherent to the surface of the brain. As shown here , the pia dips into the complex sulci and fissures that characterize the surface of the hemisphere, whereas the arachnoid does not. Since these two membranes form the borders of the subarachnoid space, small pools of CSF accumulate in these regions.

FIGURE 1-12
Two key structures cross the subdural space to enter the superior sagittal sinus. One is the constricted passage-way through which CSF must pass to reach the interior of arachnoid granulations. The second is a group of bridging veins that empty blood from the surface of the hemisphere into the venous sinus. However if, in an accident, the skull strikes a hard surface (the dashboard of a car), the brain may shift position slightly within the cranial cavity and as a result the bridging veins may be torn. These veins pass through the subdural space, and that's where the blood accumulates. Since the the dura and arachnoid easily separate, the blood in this case spreads easily over the surface of the hemisphere.

FIGURE 1-13
Here, the dura has been removed and the arachnoid, a delicate, filmy layer, can be seen lying on the surface of the brain like a wet sheet. When the dura was pulled away, the arachnoid granulations "popped out" of the superior sagittal sinus and can be seen on the left. In life, the space just underneath the arachnoid (the subarachnoid space) is filled with cerebrospinal fluid (CSF) which lifts the arachnoid off the surface of the brain and presses it against the dura.

FIGURE 1-14
This is an enlarged view of the superior sagittal sinus, taken from Figure 1-16. Since the cut was made exactly in the midline, it has passed through the middle of the sinus - we are inside the sinus looking at the lateral wall, so to speak. The little round balls within the sinus are arachnoid granulations. Admittedly, they are a little hard to see.

FIGURE 1-15
This lateral view simply shows the 6 bones that contribute to the surface of the cranial cavity. In some regions the meningeal layer of the dura breaks away from the dural one to form tough, unyielding folds or partitions which partially subdivide the cranial cavity. Two of these - the falx cerebri and the tentorium cerebelli - are shown here. These partitions are important because when expanding lesions in the cranial cavity - blood or a tumor - displace brain tissue, it is often obliged to herniate around one of them.
When the dural layers separate, as described above, they often enclose a network of venous sinuses. This is shown most clearly in this view, where the superior sagittal sinus. the inferior sagittal sinus and the superior petrosal sinus are labeled.

FIGURE 1-16
The main point of this view is to show you what some of these structures look like in "real life". Start by confirming the identity of the superior sagittal sinus, the inferior sagittal sinus, the straight sinus, the falx cerebri and he falx cerebelli.
Then, just for fun, compare the position of the lamina terminalis here and in Figure 1-8 and see if you can describe how this most rostral structure (in Figure 1-8) occupies a more central position in the adult brain.
Finally, consider this: you are working in the emergency room and someone comes in who has received a bad blow to the forehead. You notice that he seems to have a "drippy nose". You ask him if he has a cold and he says "no". What do you think of next? The clue is in this view.

FIGURE 1-17
The dura, shown in Figure 1-7, was subsequently removed from the surface of the brain and cut into these two pieces. Then india ink was injected into the middle meningeal artery. Note how the branches of the vessel stand out on the outer ( = periosteal ) surface. If you rubbed your finger over it, you would easily feel these ridges. In contrast, the inner ( = meningeal ) surface is quite smooth. When the brain moves slightly within the skull (when you bump your head against a wall, for example) the brain's surface moves, relative to the inner surface of the dura, and it makes sense for the latter to be smooth. The result of this asymmetrical split is that the artery erodes a "valley" in the inner surface of the skull., as shown in Figure 18. If the skull fracture and the fracture line crosses this valley, then there is a good chance the middle meningeal artery will be torn and that arterial blood will leak forming a hematoma (pool of blood) between the dura and the skull.
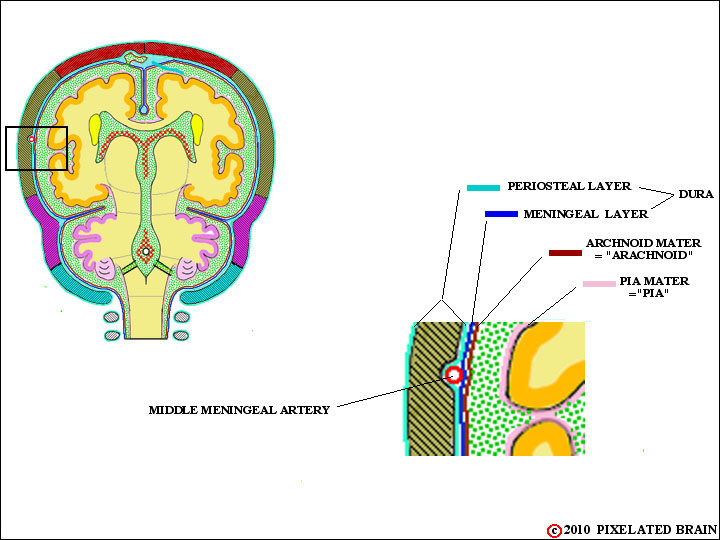
FIGURE 1-18
The layers of the dura also split to enclose the middle meningeal artery. The split is asymmetrical in that the meningeal surface - the one that the brain rubs against when it moves slightly within the cranial cavity - remains smooth, but the vessel creates a ridge in the periostial surface. The result of this asymmetrical split is that the artery erodes a "valley" in the inner surface of the skull., as shown here. If the skull fracture and the fracture line crosses this valley, then there is a good chance the middle meningeal artery will be torn and that arterial blood will leak forming a hematoma (pool of blood) between the dura and the skull.

FIGURE 1-19
At the Foramen Magnum the periostial and meningeal layers of the dura part ways (arrow A). The periostial layer continues on to the external surface of the skull, while the meningeal layer forms a tube, hanging rather freely within the vertebral canal, enclosing the spinal cord. Since the cranial dura is tightly adherent to the skull, there is - normally - no epidural space in the cranial cavity. In contrast, there is a true spinal epidural space and, as shown here, it is filled with fatty tissue and a plexus of veins. The venous plexus, known as Batson's plexus, is important because it extends the length of the vertebral canal and the veins have no valves, allowing blood flow in either direction. This forms an important route by which prostatic cancer cell may easily spread to vertebrae.
The dura forms a pressure tight seal around everything that passes through it - blood vessels and nerves (shown here) - with the result that the pressure within the cranial cavity and spinal dural sleeve differs from atmospheric pressure.

FIGURE 1-20
This view shows, in semi-schematic form, the transition from cranial to spinal dura. As Figure 19 demonstrates more clearly, there is a true spinal epidural space, filled with fat an a plexus of veins.

FIGURE 1-21
The dura is a pain-sensitive structure - but the arachnoid, pia and the brain itself are not. The dura of the anterior and middle fossae receives sensory endings from the trigeminal nerve (Figure 1-21). The dura of posterior cranial fossa is supplied by branches of the vagus nerve, coming off before it exits from the cranial cavity) and by branches of spinal nerves C2 and C3, that enter through the Foramen Magnum. The significance of this is that irritation of the dura may be perceived (or "referred" to) as coming from sensory territory of the trigeminal or upper cervical nerves.

FIGURE 1-22
In most regions the pia and arachnoid are separated by only a few mm, but in some places the gap becomes much larger and accumulations of CSF, termed cisterns, are formed in the expanded subarachnoid space. Sometimes this happens because the arachnoid fails to follow the pia into cul de sacs formed by the brain's surface; the cisterna magna, superior cistern and interpeduncular cistern are good examples. Another cistern, the prepontine one, occurs because the large basilar artery, running in the subarachnoid space, pushes the arachnoid and pia apart, as suggested in Figure 1-16.

FIGURE 1-23
Module 1 covers the anatomy of the Forebrain. Thus, as shown here, this module focuses on zone above the red line, which includes the Diencephalon and the Telencephalon (= cerebral hemisphere).
Module 2 covers the anatomy of the Brainstem and Spinal Cord.
While this seems like a very tidy division, the situation is, in fact, more complex. The Diencephalon does indeed have a close developmental and functional relationship with the telencephalon. But it is also the "gateway" to the hemisphere, in that almost all ascending sensory pathways pass through the Diencephalon to reach the cerebral cortex. So it is inevitable that we will often think of it as being the rostral end of the brainstem, and most of the discussion of it will be found in Module 2. The thalamus is by far the largest subdivision of the Diencephalon and we may at times (incorrectly) use the two terms interchangeably.

FIGURE 1-24
The meninges have been removed and we now look at the lateral surface of the brain. In this view we see the cerebral hemisphere, the brainstem and the cerebellum, but for now we will focus on the hemisphere. Clearly, there has to be a way to refer to different parts of this complex structure. We describe how it is done in the next few figures.

FIGURE 1-25
The starting point is to divide the hemisphere into four major lobes, which correspond roughly in name and position to the bones of the overlying skull.

FIGURE 1-26
This view shows the general position of the four lobes present on the lateral surface of the hemisphere.

FIGURE 1-27
The landmarks used to establish the borders of each lobe are shown here. The central sulcus lies about halfway between frontal and occipital poles and is significantly deeper than adjacent sulci. The lateral fissure differs from a sulcus in that it was created as tissue migrated in an arc-like fashion to form the temporal lobe. The pre occipital notch is a rather predictable indentation in the surface of the brain, and the parieto-occipital sulcus is a very constant and deep sulcus, present on the medial surface of the hemisphere. The anterior border of the occipital lobe is a line drawn between the pre occipital notch and the upper end of the parieto-occipital sulcus.

FIGURE 1-28
Anteriorly, the lateral fissure separates the temporal lobe from the frontal and parietal lobes; posteriorly, the line separating the temporal and parietal lobes is a somewhat arbitrary extension of the lateral fissure. Because of their unique geometry the insula and the limbic "lobe" to not fit into the four lobe scheme. As the name suggests, the insula is indeed an island of cortex that has been buried by the expanding frontal, parietal and temporal lobes; it shares borders with all three. The limbic "lobe" runs around the medial edge of the hemisphere, sharing borders with all four lobes. Because of its functional importance it is commonly called a lobe, though it hardly seems like one.

FIGURE 1-29
Here are some of these landmarks on a real brain.

FIGURE 1-30
Before describing how each lobe is subdivided into gyri, we need to be clear on the way directions are specified when dealing with the surface of the hemisphere. The most commonly used terms are shown in this view.
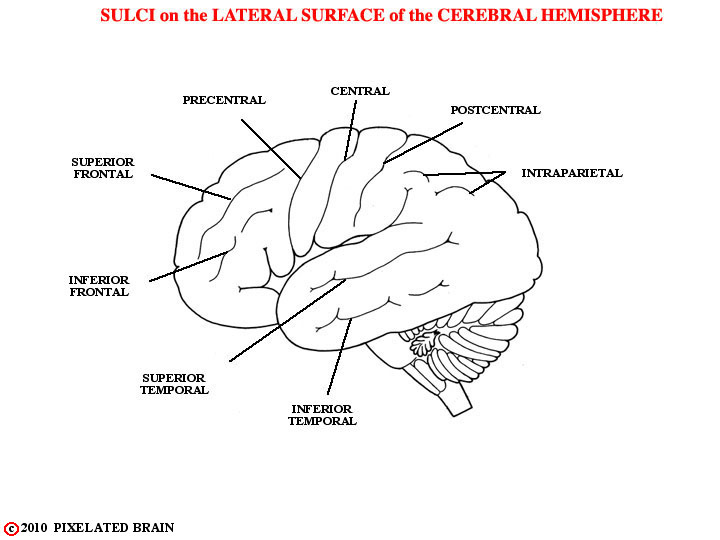
FIGURE 1-31
These are the named sulci.

FIGURE 1-32
These are the named gyri.

FIGURE 1-33
Most textbook figures imply that the pattern of gyri and sulci on the surface of the hemisphere is quite similar from one brain to the next. In reality, there is a great deal of variability between specimens. In the brain we looked at originally, for example, the central, post central and superior temporal sulci are rather clear, with the result that the post central gyrus and the superior temporal gyrus are easily identified, as shown here. In other brains, even the central sulcus (the most obvious of the three) can be difficult to identify.

FIGURE 34
Migration of the frontal, parietal and temporal lobes has tended to bury an "island" of cortex within the depths of the lateral fissure (Figure 1-39 , Figure 1-40) . If parts of the frontal (red) and parietal (yellow) lobes are cut away this cortical area - the insula (violet) - is revealed, as shown here. Blumenfeld shows this in his Figure 2.24. The insula is not truly an island, however, and its relationship to the lobes of the hemisphere is best revealed by making a cut through the brain in the plane indicated by the line and looking at the surface of the sectioned tissue. We will see the section in the next view.

FIGURE 1-35
The insula is not truly an island. Its relationship to the lobes of the hemisphere is best revealed in a section such as this one.

FIGURE 1-36
The anatomy in this basal view becomes a little more complex, because we see not only the basal surface of the hemisphere, but also the surface of the brainstem - the complex region lying between the hemisphere and the spinal cord, and the subject of Module 2. For now, we will focus exclusively on the hemisphere. In this view. we identify the lobes of the hemisphere.

FIGURE 1-37
Here, we label the sulci and gyri. The uncus is an important landmark on the medial edge of the temporal lobes.

FIGURE 1-38
The medial surface of the hemisected brain is the most interesting of all because it reveals not only another aspect of the hemisphere but also the midsagittal surfaces of the diencephalon and brainstem. We will leave the brainstem structures for Module 2 and mainly look at the hemisphere.

FIGURE 1-39
These are the gyri seen on the medial surface of the hemisphere.

FIGURE 1-40
More anatomy.

FIGURE 1-41
At this point, you are asking "Is that it? Do I have all the names I need to describe the surface of the cerebral cortex? If so, now tell me what each of these gyri actually does." Well, the gyri are a necessary start, but its not that simple (it never is). The gyri are useful landmarks for the neurosurgeon, the neuroradiologist and the neuropathologist and they are used in the reports that these specialists write - operative notes, and the like. But it turns out that the cellular region lying just deep to the surface of the hemisphere - the light area in Figure 1-35 - is organized into 6 layers and the thickness and cellular structure of these layers differs from one region to another. More than 100 years ago, a worker by the name of Brodman parcelled the surface of the cortex out into more than 50 areas, based on the destinctive cellular appearance of each, and his map, shown here, is used to this day in describing the functional organization of this region.

FIGURE 1-42

FIGURE 1-43

FIGURE 1-44
Migration of the frontal, parietal and temporal lobes has tended to bury an "island" of cortex within the depths of the lateral fissure, as seen here and in the next figure.